Número 017 2004 Revista Internacional de Psicoanálisis Aperturas
Estudios sobre la relación herencia-ambiente en la temprana infancia
Autor: Dio Bleichmar, EmilcePalabras clave
Co-accion, Factores ambientales, Factores geneticos, Factor temporal de la interaccion, Regulacion bidireccional de la conducta, Significado subjetivo de la experiencia..
Versión ampliada de la conferencia pronunciada en el Congreso Internacional "Herencia y ambiente en psiquiatría del niño y del adolescente". Hospital General Universitario Gregorio Marañón, Madrid (Mayo 2004)
¿Genética de la conducta versus parentalización?
Cuánto juegan el código genético por un lado y la influencia del ambiente, por el otro, en moldear el desarrollo del cerebro es una de las cuestiones que ha polarizado a los científicos. Las posiciones han estado determinadas más por preferencias ideológicas que por sólidos datos que permitiesen estudiar la complejidad de la influencia de uno y otro de los factores. A lo largo de la última década del siglo XX, provocado tal vez por la excitación del proyecto del genoma humano pero, también, por los diseños de investigación con una sofisticación estadística cada vez mayor, la investigación evolutiva ha estado dominada por ideas provenientes de la genética cuantitativa de
Afortunadamente, esta reflexión -que podría entenderse como un comentario más pero sin fundamento- empieza a cobrar peso ya que la experimentación en animales comienza a demostrar la necesidad de contar con modelos relacionales, interactivos, de causalidad en que los factores genéticos y ambientales aparecen operando en conjunción.
Las investigaciones actuales muestran al cerebro como operando de forma plástica y autoorganizada y estando menos constreñido por límites predeterminados de lo que se había pensado previamente. La información en el cerebro es representada y procesada por grupos de neuronas que mantienen una interconexión funcional basada más en las exigencias de la experiencia que en estrictos esquemas genéticamente determinados.
La comprensión de los fenómenos del desarrollo requiere conceptos relacionales o co-activos de la causalidad en oposición a causas únicas que operan en un supuesto aislamiento (Gottlieb, G. & Tucker Halpern, C. ,2002). El concepto llave es entender que lo que hace que el desarrollo suceda es la relación entre los dos componentes, y no los componentes en sí (persona-persona, organismo-organismo, organismo-ambiente, célula-célula, gen-gen. actividad-conducta motora). Cuando se habla de co-acción como el corazón de la causalidad evolutiva lo que se quiere enfatizar es que necesitamos especificar alguna relación entre al menos dos componentes del sistema del desarrollo. El concepto usado más frecuentemente para designar la co-acción es el relativo a la experiencia.
Voy a mencionar en primer lugar algunos trabajos sobre la influencia del medio en la modificación del cerebro en animales en períodos críticos y en segundo término estudios en la relación temprana en humanos que van en la misma línea.
Influencia del medio externo en la modificación del cerebro
En el nº 404 de
Mriganka Sur y colaboradores (Massachussets Institute of Technology, USA) han perfeccionado una técnica que les permite redirigir quirúrgicamente hacia qué parte de la corteza cerebral se dirigirán los nervios en el roedor hurón de un día. Más específicamente, las conexiones de la retina que normalmente llegan a la corteza visual son redireccionadas para mandarlas hacia la corteza auditiva. Esto determina que la corteza que estaba genéticamente preparada para ser sensible a los estímulos auditivos pase a ser capaz de captar y de organizar percepciones visuales.
Eso no es todo: la corteza de la región que hubiera tenido una estructura anatómica de corteza auditiva pasa a poseer una disposición de sus células correspondientes a la corteza visual (con una organización neuronal -ubicación en molinillo- que le permite convertirse en un mapa de las distintas zonas de la retina y de la sensibilidad de ésta frente a formas en el espacio). Pero los experimentos de Sur y col. van más allá de contentarse con probar que la corteza preparada genéticamente para estar organizada con cierto tipo de disposición celular adquiere los caracteres de la corteza visual al recibir los estímulos visuales: el animal, una vez crecido, pasa a ser capaz de responder conductualmente a los estímulos visuales en la corteza que genéticamente estaba predeterminada para ser área auditiva.
Es decir, si la corteza cerebral de una determinada zona recibe estímulos de un tipo dado, son estos estímulos los que ocasionan la organización y la funcionalidad que tendrá y no la predisposición genética. En otras palabras, es la naturaleza del estímulo externo el factor decisivo de cómo se terminará organizando una cierta zona cortical.
Cierpial & Mc Carty (1987) encontraron que ratas con hipertensión espontánea (SHR) usadas como modelo animal para el estudio de la hipertensión humana se vuelven hipertensas por co-acción con las madres después del nacimiento. Cuando las ratitas SHR son amamantadas y criadas por madres normales desde el nacimiento no desarrollan hipertensión. Pareciera que hay un componente hiperactivo en la conducta de las madres que induce a las ratitas SHR a desarrollar hipertensión. La naturaleza interactiva de la articulación entre gen y crianza del desarrollo de la hipertensión en ratas SHR es demostrada por el hecho que ratas que genéticamente son normotensas no desarrollan hipertensión cuando son amamantadas y criadas por madres que son genéticamente hipertensas (SHR). De modo que aunque las ratitas genéticamente hipertensas (SHR) difieren de las normales, el desarrollo de la hipertensión requiere la co-acción con las madres hiperactivas, no es resultado inevitable del hecho que son genética, anatómica y fisiológicamente diferentes de las normales. Es un buen ejemplo de los aspectos relacionales de la definición de la experiencia y de la causalidad del desarrollo. La causa de la hipertensión no reside en la genética de las ratas SHR o en las de las madres SHR sino en la relación que se establece entre ellas.
La breve separación de sus madres en el período previo al destete produce en ratitas cambios en el eje hipotálamo-hipófisis-adrenal en las respuestas a estímulos estresantes (Plotsky & Meaney 1993). Cabib et al. (2000) demostraron que la respuesta a las anfetaminas puede ser alterada dramáticamente en ratas adultas ante un acontecimiento como la escasez de alimento, un cambio mediado por la secreción de glucocorticoides y liberación de dopamina. En la discusión sobre la inevitable interacción entre diferencias genéticas y factores ambientales estresantes, estos investigadores explícitamente señalan que "el intento de identificar una causalidad genética o ambiental como factores independientes es probablemente lógica pero procedimentalmente errónea" (p. 464-465).
Una de las dimensiones humanas que goza de mayor crédito genético es
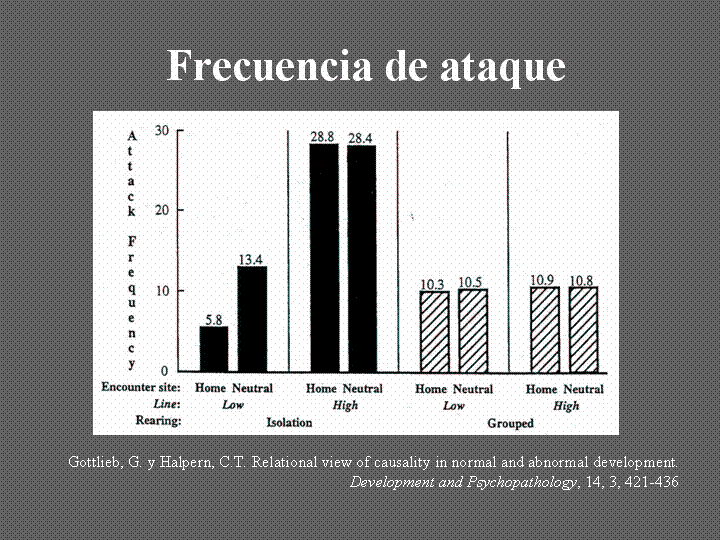
Hallazgos todos que apoyan la idea que es la co-acción de estrés, trauma y otro tipo de factores patógenos externos junto con configuraciones genéticas lo que en última instancia da como resultado diferentes perfiles patológicos. Se subraya el punto que ni un genotipo particular ni un factor traumático externo están inevitablemente vinculados a un resultado patológico. Es la particular combinación de factores genéticos y ambientales y probablemente el factor temporal de la interacción que provoca resultados patológicos.
Con todo, se podría argumentar que esta plasticidad cerebral se da sólo en roedores, que no tiene nada que ver con lo que sucede en humanos. Sin embargo, Sadato (1996) y Rauschecker (1995) ya habían mostrado cómo en sujetos ciegos desde muy temprano la corteza visual pasaba a responder a estímulos táctiles, siendo activada, por ejemplo, cuando se leía en sistema Braille. Estos experimentos dieron lugar a que un equipo integrado por investigadores del National Institute of Health (USA) y de la Facultad de Medicina de Fukui (Japón) desearan poner a prueba la hipótesis de que la plasticidad cerebral -cambios producidos por una actividad específica- requería que se hubiera producido en una época muy temprana de la vida, es decir, en aquellas personas que eran ciegas de nacimiento o poco después del nacimiento. Repitieron los experimentos tendentes a ver si la corteza visual en ciegos después de los 14 años respondía a estímulos tactiles al leer en Braille y constataron que esa capacidad de la corteza occipital (visual) de reconocer estímulos táctiles ya no existía cuando la ceguera había ocurrido después de esa edad.
Los hallazgos conducen a la idea cada vez más fundamentada que en humanos hay un período en que el cerebro es plástico, durante el cual se organiza la funcionalidad y que, superada esa "ventana" durante la cual el cerebro se puede modificar en una dirección determinada por el estímulo externo, después ya resulta mucho menos factible.
El efecto del niño sobre los padres
Los estudios sobre gemelos realizados en los Estados Unidos, Escandinavia, el Reino Unido y otros lugares se han tomado como punto de referencia y a partir de ellos se ha extendido la idea que los genes son más importantes que el entorno en casi todas las enfermedades psicológicas. Gemelos idénticos que habían sido criados por separado revelaron semejanzas llamativas en sus conductas, personalidad y formas de pensar (Reiss y col., 1995; Neubauer, 1996; Plomin y col., 1997; Reiss y col., 2000). Es bastante probable que pueda haberse exagerado la importancia de los padres para el desarrollo: los estudios sobre adopción, concretamente, muestran que mucha de esa influencia parental es ilusoria. Es también probable que las características de personalidad del niño que se han considerado como reacción a la conducta parental sean en muchos casos predisposiciones genéticas y que el rasgo de la personalidad y la forma de parentalidad -crítica, cálida o incluso abusadora- sean consecuencias de los mismos genes en los padres y en el niño. Del mismo modo, los estudios sobre adopción sugieren que los niños con tendencias genéticas hacia la agresión provocarán una parentalidad más hostil y coercitiva (Ge y col., 1996).
De modo que, si bien los estudios sobre adopción y gemelos demuestran que el trastorno psicológico tiene raíces genéticas, los efectos genéticos son en su mayor parte indirectos más que directos. Incluso una carga genética alta para un riesgo ambiental no significa que el resultado asociado se transmita genéticamente de forma necesaria. Por ejemplo, aun en el caso que se hallara que un abuso infantil y sus secuelas tienen base genética, el trastorno de un niño que ha sufrido abusos seguiría comprendiéndose mejor en términos de la destrucción de la confianza que el niño tiene en el mundo. La pérdida de confianza es la que tendría que encararse en cualquier intervención terapéutica. Los datos conductuales genéticos no nos ayudan demasiado en la situación clínica individual.
Los trabajos en la denominada interacción gen-ambiente en animales nos van ayudando a saber cómo funcionan los genes. Al menos parte de cada gen es un mecanismo de control para el proceso de transcripción, en otras palabras, determina si un gen se expresará o no en ciertos rasgos observables físicos o psíquicos. Las experiencias internas y externas, las hormonas, el estrés, el aprendizaje y la interacción social alteran la unión de los reguladores de transcripción (Kandel, 1998). Por ejemplo, las crías de ratas separadas de sus madres en las dos primeras semanas de vida muestran un incremento permanente en la expresión (manifestación en rasgos) de genes encargados de controlar la secreción de FLC (factor liberador de corticotropina), una de las hormonas del estrés (Plotsky y Meaney, 1997). Sin embargo, esta vulnerabilidad al estrés, que dura toda la vida, se invierte si las madres demuestran un mayor cuidado físico a las crías una vez que se las reúne (Liu y col., 1997).
De modo que la cuestión es saber cuáles de los aspectos del entorno influyen en estos mecanismos de control en los humanos. Existen unos pocos ejemplos importantes en este sentido, pero aislados. Los hijos de padres/madres esquizofrénicos que son adoptados lejos de ellos desarrollan la enfermedad si su familia adoptiva es disfuncional (Tienari y col., 1994). Se han hecho observaciones similares en cuanto a la conducta criminal (Bohman, 1996). De modo que el riesgo genético puede hacerse realidad o no, dependiendo del entorno familiar.
Pero los genetistas conductuales se encuentran con un problema. A pesar de su aceptación de un modelo de interacción gen-entorno, ha habido pocos hallazgos que demuestren el efecto moderador del entorno en la expresión del gen en los humanos (Plomin, DeFries y col., 1997). La genética de la conducta humana estudia en su mayor parte el entorno objetivo, externo, familiar. El entorno que estimula la expresión de un gen no es objetivo, no es observable. La distinción de Freud entre las dos superficies de la conciencia -una vuelta hacia dentro y otra vuelta hacia fuera- proporciona la pista: es la experiencia del entorno lo que produce interacciones entre la herencia y la circunstancia, no el hecho de esa circunstancia en sí misma. La interacción es entre el gen y el entorno subjetivo.
El modo en que se experimenta al entorno actúa como filtro en la expresión del genotipo en el fenotipo, la traducción del potencial genético en personalidad y conducta. Como sostiene Fonagy, aquí tocamos la importancia del psicoanálisis para la comprensión de las influencias genéticas sobre
El que un factor ambiental desencadene o no la expresión de un gen puede depender del modo en que el individuo interprete esa experiencia, determinada a su vez por significados conscientes o inconscientes atribuidos a la misma (Kandel, 1998). Así, los procesos representacionales intrapsíquicos no son sólo consecuencia de los efectos ambientales y genéticos, sino que es probable que sean moduladores vitales de dichos efectos. El ganador del Premio Nobel del 2000, Eric Kandel, llegó a sugerir que los cambios más profundos y a más largo plazo asociados con la terapia psicoanalítica pueden sobrevenir mediante los cambios en la expresión genética provocados por las transformaciones intrapsíquicas que se operan en el transcurso de una psicoterapia que apunte a la subjetividad consciente e inconsciente.
En un programa de tratamiento psicoanalítico preescolar en
Fonagy (2003) sostiene que incrementar la conciencia emocional, como lo hace la psicoterapia exitosa, podría haber reducido la expresión de esta vulnerabilidad genética a los problemas psiquiátricos adultos mediante la elaboración de la representación mental de las relaciones intersubjetivas. Enfatiza que se trata de resultados muy preliminares pero lo que quiere resaltar se refiere a una cuestión conceptual: que los genetistas conductuales, los que toman en cuenta sólo acontecimientos, tendrán más dificultadoes para otorgar un sentido a los datos que recojan sobre la mente a menos que incluyan en sus estudios sobre el entorno o ambiente el significado subjetivo –consciente e inconsciente- que poseen los acontecimientos, especialmente aquellos que, una vez realizadas ciertas interpretaciones, podrían tener un impacto emocional importante desencadenando ansiedad o depresión sostenidas.
Importancia de la intersubjetividad y de la capacidad reflexiva en los períodos críticos de plasticidad cerebral
Si algo tan rígidamente predeterminado como la corteza cerebral depende para su desarrollo anatómico y funcional del estímulo externo, y existen períodos críticos para que esta influencia se ejerza, ello aporta datos coincidentes con la importancia que en psicoanálisis se otorga al desarrollo de los sistemas motivacionales -apego, sensual/sexual, narcisista, autoconservación, regulación psicobiológica- en los períodos iniciales de la vida y, especialmente, a los estímulos de las figuras significativas en moldearlos. En el interjuego entre lo genéticamente determinado -el instinto- y la influencia de las figuras externas -la realidad-, éstas últimas resultan trascendentes para reorientar lo instintivo en una dirección u otra. Es aquí donde el concepto psicoanalítico de pulsión/motivación como algo diferente del instinto adquiere su valor. La pulsión será el principio motivacional organizador de la conducta, con igual fuerza que un instinto, incluso con la perentoriedad con la que se sigue rígidamente un esquema determinado de acción, pero que ya no es el instinto simplemente predeterminado por el programa genético sino la reorientación que la experiencia, en los intercambios con las figuras significativas, le imprime a éste.
La capacidad para la interpretación, que el filósofo Bogdan (1997) ha definido recientemente como “organismos que se otorgan sentido entre sí en contextos donde esto tiene importancia biológicamente”, es una característica de todas las especies complejas y se convierte en algo únicamente humano cuando la materia de la interpretación pertenece al dominio de los estados mentales. Esta capacidad es la que proporcionaría el paso final en la trascripción de la influencia genética en un patrón de conducta. El mecanismo interpretativo codifica la información genética en forma de tendencias, pero también modera la influencia genética modificando la percepción que el niño tiene de su mundo de objetos.
Sabemos que este modulador entre genotipo y fenotipo es una capacidad que depende de forma crucial de la respuesta sensible de los cuidadores tempranos a los estados emocionales del bebé. Una función vital de la relación temprana -que se suele sumar al concepto de apego cuando en realidad lo sobrepasa- es el contexto que la relación infante-madre provee para la adquisición de la comprensión de los estados mentales: los componentes básicos de la autoorganización y
Existen estudios que relacionan déficits concretos en la sensibilidad parental con formas de psi
Gergely y Watson (1996), analizando en una microestructura relacional la así llamada sensibilidad materna sugieren contundentemente que la respuesta parental sensible implica en la práctica dos parámetros: uno de correspondencia o contingencia entre lo que siente el niño y lo que responde emocionalmente la madre; el otro parámetro, es la comunicación de la diferencia entre lo que siente la madre y el niño. Acuñan el término marcación para designar al hecho de que cuando una madre está especularizando acciones del infante, las acciones maternas que comunican al infante tienen la cualidad de que lo que está mostrando no es su propio sentimiento sino una representación de la percepción que ella tiene de la experiencia del infante -por ejemplo, cuando el niño siente dolor, la madre puede decir con un rostro que simula parte de la expresión del bebe pero acompañado de un "¡pobrecito, cómo te duele!", dicho con tono cariñoso no sufriente. Las madres que pueden “marcar o señalar” su expresión emocional añaden así un conjunto especial de atributos a la expresión del afecto del niño que las diferencia claramente de su propia expresión de afecto, y esto parece ser el factor clave que permite consolar al infante con bastante más rapidez. El infante capta que el estado emocional que la madre muestra tiene relación con el que él siente y no que la madre sienta exactamente lo mismo que él.
Los autores sugieren que la especularización del afecto por parte del adulto que refleja el sentimiento del infante pero carece de esta marcación genera problemas evolutivos dentro del espectro borderline. Este patrón de especularización puede esperarse en madres que, debido a sus propias dificultades con la regulación emocional, se ven abrumadas por la angustia de sus infantes. Puesto que la especularización del afecto no está marcada, ésta no se desconectará del cuidador y se percibirá como correspondiente a una emoción parental real. El infante sentirá que su estado emocional es más peligroso y atemorizante puesto que parece contagioso. Dado que el infante atribuye el afecto especularizado a la figura parental, sentirá que su propia angustia “está ahí fuera”, que pertenece al otro más que a sí mismo. A corto plazo, la percepción de la angustia correspondiente en la figura parental no regulará sino que aumentará el estado negativo del bebé, pudiendo dar lugar a la traumatización más que a la contención emocional.
Otro tipo importante de estructura especularizante desviada se produciría por la dominancia de una especularización marcada pero incongruente. Pensemos en un infante cuya excitación erótica por el contacto físico induce en la madre angustia y enojo defensivo debido a sus conflictos en relación con
El significado subjetivo en la transmisión intergeneracional
Los estudios de la regulación afectiva del bebé por parte del adulto, incluida su reactividad neurovegetativa, no dejan lugar a dudas acerca de que las interacciones tempranas moldean, modulan dimensiones que tienen un claro componente genético. Esto sucede también con las modalidades de apego para las cuales se ha evidenciado la importancia de componentes cerebrales innatos (Insel, 1997; Leckman y col., 2004).
Uno de los mayores méritos de la Entrevista de Apego para Adultos desarrollada por Mary Main (AAI) -el procedimiento que permitió a la teoría del apego dar un giro desde lo puramente etológico y conductual hacia el nivel representacional- fue el de mostrar cómo un tipo de apego del adulto tiene una alta probabilidad de reproducirse en la siguiente generación. La entrevista hecha a la madre (un cuestionario sobre su propia experiencia de apego) se ha evidenciado como un excelente procedimiento para la predicción del sistema de cuidado que luego ésta empleará en la crianza del niño. Esta correlación es la que ha permitido entrever los mecanismos de la transmisión intergeneracional y ha sido empleada por numerosos investigadores en distintos países (Hesse, 1999). En el estudio de Fonagy y col. (1991) se mostró que la AAI aplicada a mujeres embarazadas tiene una alta capacidad predictiva del tipo de apego que presentará su hijo/a a los 12 y 18 meses de edad. Madres que antes del nacimiento de su hijo manifestaron desapego en la narración de su propia infancia, tendieron en su mayor parte a tener bebés que se comportaban hacia ellas de forma notoriamente evitativa al reencontrarse en la condición experimental conocida como situación extraña. Los valores que aportan estos autores son muy impresionantes: un 75 % de concordancia entre lo que se detecta en la entrevista con la madre sobre cómo fue su apego y el tipo de apego que mostrarán luego sus hijos. Encontraron también relación entre la seguridad del apego del infante al padre a los 18 meses y la entrevista realizada al padre antes del nacimiento del hijo
A diferencia de la clasificación sobre los patrones de apego en el infante, a quien se lo considera seguro o inseguro con respecto a uno de los padres, en particular con quien se lo está observando (un infante puede mostrar un apego seguro con la madre y evitativo con el padre), la ubicación del adulto dentro de la categoría seguridad en la Entrevista de Apego para Adultos no se identifica con ninguna relación en particular. No se evalúa el apego de un adulto con relación a ninguna figura, ni del pasado ni del presente, y por lo tanto el análisis de la entrevista no permite clasificar al adulto como teniendo un apego seguro o inseguro. La entrevista del adulto con el instrumento AAI, aun sin familiares vivos o después de un amargo divorcio, puede ser clasificada como segura/autónoma ya que lo que se codifica son estados de la mente con respecto a su historia global de los vínculos de apego, tal como se manifiesta en el contexto de la entrevista.
Como queda demostrado en la prueba de la Entrevista de Apego para Adultos, lo importante en una relación no es tanto el grado de sufrimiento o frustración vivido sino si este hecho se constituye en muro de piedra emocional, retenido como gesto o memoria corporal, o ha pasado a un registro simbólico que permite su circulación y transformación. De ahí que nos interese discernir en la medida de lo posible las modalidades fantasmáticas a partir de las cuales la madre ha dado significado a los acontecimientos de la crianza.
De acuerdo a los resultados de la aplicación de la prueba los padres pueden ser clasificados habiendo tenido una historia de apego: a) segura/autónoma; b) desentendida o despreocupada (no se enteran o no le dan importancia a los temas de apego); c) preocupada. La valoración como apego seguro no depende de la normalidad y ausencia de trastornos psicológicos del enunciante y de las vicisitudes de su infancia sino de la coherencia, autenticidad emocional y comprensión psicológica de su relato sobre la relación con sus padres. Puede ser un adulto que haya pasado situaciones traumáticas y no obstante ser categorizado como seguro/autónomo. Otro ejemplo ilustrativo de que la valoración no depende de factores objetivos, sino de la capacidad del sujeto de otorgar sentido y comprender contextualmente lo sucedido.
De modo que la predicción del tipo de cuidado que, de acuerdo a la AAI, se dará al infante va a depender no de la historia de la vida de la madre en sí sino de la forma en que es contada, o sea, cómo ha sido procesada la experiencia, y no la experiencia en sí misma. Mientras la historia de la vida de un individuo no puede cambiar, no obstante puede ser contada o reconstruida en muchas formas diferentes y este procesamiento diferente constituye la capacidad de modificación del destino.
La regulación bidireccional de la conducta
Estudios empíricos sobre microinteracciones cara a cara en la díada madre-infante muestran cómo la acción y la información es conjuntamente construida, esto quiere decir, regulada bidireccionalmente (Beebe y col. 1997). La experiencia se realizó con madres y bebés de 4 meses filmados en dos tipos de observaciones:
a) la regulación bidireccional del reflejo facial;
b) estructuras de interacción de descarrilamiento (Chase-and-dodge)
Para el reflejo facial la escala está basada en la orientación, la mirada y las expresiones faciales. Por la velocidad del cambio se deduce que antes que el comportamiento del compañero esté terminado, el otro está empezando a hacerlo, de modo que ya se han creado esquemas anticipatorios faciales-visuales (la duración de los comportamientos está en el rango de 1/4 a 1/3 a 1/2 de segundos). El reflejar facialmente la cara del otro se considera un precursor de la empatía y una de las estructuras de interacción que contribuye a la organización presimbólica de las representaciones del sí mismo y del otro.
Ekman (1983) y Zajonc (1985) muestran que en el entonamiento emocional la expresión del otro está altamente correlacionada con el patrón de activación fisiológica. Una expresión particular de la cara está asociada a un modo particular de actividad autonómica, y reproduciendo la expresión de otra persona se produce un estado fisiológico similar en el espectador. Tronick (1989) aporta datos demostrando que en el transcurso de una interacción exitosa madre-hijo se corresponden sólo aproximadamente un 30% del tiempo, el resto del tiempo pueden estar ligeramente desentonados.
La segunda parte del estudio de Beebe encara las estructuras de ruptura y reparación del entonamiento interactivo. Cada madre puede sobrepasar así como no llegar a un óptimo nivel de estimulación. La instruccion que se le da a la madre es: "juega con tu bebé como lo harías en casa". La estructura de la interacción documentada a través de secuencias estadísticamente significativas es como sigue: mientras la madre se acerca a la cabeza del bebé, el bebé se mueve hacia atrás y hacia fuera. La madre después "persigue" moviendo su cabeza y su cuerpo hacia el bebé. Al mismo tiempo que le persigue, el bebé simultáneamente mueve la cabeza hacia fuera todavía más lejos. Estos ajustes mutuos son semisincronizados.
El bebé tiene poder de "veto", él puede impedir totalmente un encuentro visual con la madre. Aunque ésta es una interacción aversiva, su regulación es todavía bilateral, el movimiento de la madre hacia la cara del bebé influye en el movimiento de alejamiento del bebé, y los movimientos de alejamiento de cabeza y cuerpo del bebé de la madre influyen en la persecución de ésta. Se ha llamado al juego "perseguir y esquivar" pero también podría llamarse "esquivar y perseguir". La retirada del bebé provoca la intrusión de la madre y la intrusión de la madre influye en la retirada del niño.
Los sistemas diádicos desde el punto de vista de la comunicación son la base del concepto de organización interactiva de la experiencia y de las representaciones (Beebe y col. 1992). Madre-hijo construyen conjuntamente un patrón secuencial de movimientos y reglas para regular estos movimientos a través de las dimensiones de tiempo, espacio, afecto y activación. Por repetición se crea la expectativa de un patrón característico de secuencias diádicas, junto con las consecuencias de la autorregulación que es el contenido de las representaciones del infante. Entonces lo que es inicialmente representado no es una persona, un otro, sino una relación: el sí mismo y el otro.
Lo que es representado es un fenómeno emergente de la diada que no puede ser descrito en base a cada compañero solo (Beebe y Stern, 1977). La estructura diádica inherente de estas representaciones implica que ambos roles en la interacción han sido experimentados. Esto puede explicar por qué en la vida adulta estas posiciones pueden ser intercambiables en la intimidad: masoquismo-sadismo, depredador-víctima, perseguidor-evitativo, abandonado-abandonador. Las estructuras de interacción son un código mutuamente organizado y mutuamente entendido en el cual cada rol implica su recíproco y ninguno puede ser representado sin el otro.
Conclusiones
Las comprobaciones de laboratorio van indicando una dirección en torno a la causalidad de los fenónenos del desarrollo como un sistema de influencias bidireccionales de múltiples niveles en constante interacción.
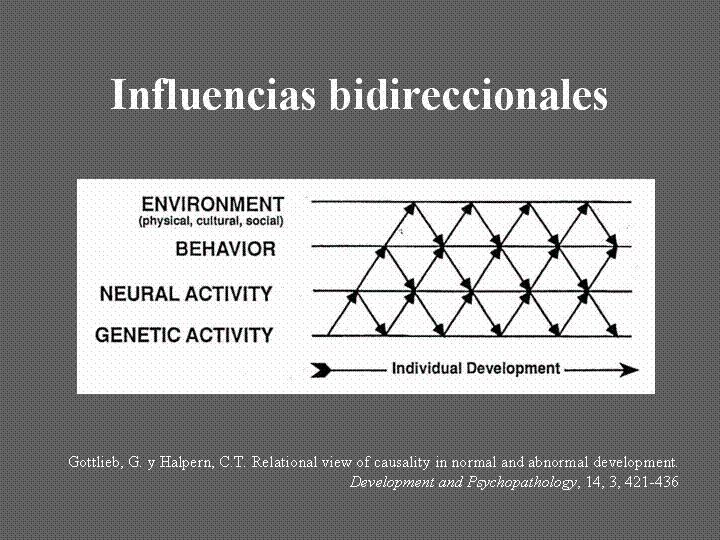
Desde hace unos años venimos trabajando en un enfoque del desarrollo psíquico al que denominamos modular-transformacional en torno a la articulación de diversos sistemas motivacionales. Modelo complejo que rompe con explicaciones dinámicas monocordes y reduccionistas de la psi
Bibliografía
Beebe, B., Lachmann, F., Jaffe, J. (1997). Mother-Infant Interaction Structures and Presymbolic Self and Object Representations. Psychoanalytic Dialogues, 7, 133-182.
Beebe, B. & Jaffe, J. & Lachman, F. (1992). A dyadic systems view of communication. In: Relational Perspectives in Psychoanalysis, ed. N. Skolnic & S. Warshaw. Hillsdale, NJ: The Analytic Press, pp. 61-81.
Beebe, B., & Stern, D. (1977). Engagement-disengagement and early object experiences. In: Communicative Structures and Psychic Structures, ed. N. Freedman & S. Grand. New York: Plenum Press, pp. 35-55.
Bleichmar, H. (2000). Influencia del medio externo en la modificación del cerebro. Aperturas Psicoanalíticas, nº 5. (Revista internacional por Internet: www.aperturas.org ).
Bleichmar, H. (1997). Avances en Psicoterapia Psicoanalítica. Barcelona. Paidós
Dio Bleichmar, E. (2000). Lo intrapsíquico y lo intersubjetivo: metodología de la psicoterapia de la relación padres-hijos/as desde el enfoque modular-transformacional. Aperturas Psicoanalíticas, nº 15. (Revista internacional por Internet: www.aperturas.org )
Bogdan, R. J. (1997). Interpreting Minds. Cambridge, MA: MIT Press.
Bohman, M. (1996). Predisposition to criminality: Swedish adoption studies in retro-spect. In: Genetics of Criminal and Antisocial Behaviour, ed. M. Rutter. Chichester, England: Wiley.
Caspi, A., McClay, J., Moffitt, T. E., Mill, J., Martin, J., Craig, I. W., Taylor, A. & Poulton, R. (2002), Role of genotype in the cycle of violence in maltreated children. Science, 297 (5582):851-854.
Cabib, S., Orsini, C., Le Moal, M., & Piazza, P.V. (2000). Abolition and reversal of strain differences in behavioral responses to drugs of abuse after a brief experience. Science, 289, 462-465.
Cierpial, M. A. & McCarty, R. (1987). Hypertension in SHR rats: Contribution of maternal environment. American Journal of Physiology, 253, 980-984.
Ekman, P. (1983). Autonomic nervous system activity distinguishes among emotions. Science, 221: 1208-1210.
Elman, J. L., Bates, A. E., Johnson, M. H., Karmiloff-Smith, A., Parisi, D. & Plunkett, K. (1996). Rethinking Innateness. Cambridge, MA: MIT Press.
Ge, X., Conger, R. D., Cadoret, R., Neiderhiser, J. & Yates, W. (1996). The develop-mental interface between nature and nurture: A mutual influence model of child antisocial behavior and parent behavior. Dev. Psychol., 32:574-589.
Fonagy, P. (2003). Genética, psi
Fonagy, P., Stein, H. & White, R. (2001). Dopamine receptor polymorphism and susceptibility to sexual, physical and psychological abuse: Preliminary results of a longitudinal study of maltreatment. Presented at 10th biannual meeting of the Society for Research in Child Development, Minneapolis, MN, April 21.
Fonagy, P., Steele, M., Steele, H., Moran, G., Higgitt, A. Target, M. (1991). La capacidad para entender los estados mentales: el Self Reflexivo en padres e hijos y su significación para la seguridad del apego. Infant Mental Health Journal (Traducción Ficha de ELIPSIS). 12, 201-218
Gergely, G. & Watson, J. (1996). The social biofeedback model of parental affect-mir-roring. Internat. J. Psycho-Anal., 77:1181-1212.
Gottlieb, G. & Tucker Halpern, C. (2002). A relational view of causality in normal and abnormal development. Development and Psychopatology, 14, 421-435.
Harris, J. R. (1998). The Nurture Assumption. New York: Free Press
Hesse, E. (1999). The Adult Attachment Interview. Historical and Current Perspective. In J. Cassidy y P. Shaver, Handbook of Attachment. New York-London. The Guilford Press
Jaffe, J., Beebe, B., Feldstein, S., Crown, C. L. & Jasnow, M. D. (2001), Rhythms of dialogue in infancy. Monogr. Soc. Res. Child Dev., 66(2).
Hood, K. E. & Cairns, R., B. (1989). A developmental-genetic analysis of aggressive behavior in mice: IV. Genotype-enrivonment interaction. Aggressive Behavior, 15, 361-380.
Insel, T. R. (1997). A neurobiological basis of social attachment. Am. J. Psychiatry, 154, 726-735.
Jaffe, J., Beebe, B., Feldstein, S., Crown, C. L. & Jasnow, M. D. (2001) Rhythms of dialogue in infancy. Monogr. Soc. Res. Child Dev., 66 (2).
Kandel, E. R. (1998). A new intellectual framework for psychiatry. Amer. J. Psychiat., 155:457-469.
--- (1999). Biology and the future of psychoanalysis: A new intellectual framework for psychiatry revisited. Amer. J. Psychiat., 156:505-524.
Leckman, J., Feldman, R., Swain, J., Eicher, V., Thompson, N., Mayes, L. (2004). Primary parental preoccupation: circuits, genes, and the crucial role of the environment. Journal of Neural Transmission, 111, p. 753-771
Lendvai, B., Stern, E. A., Chen, B. y Svoboda, K. (2000). Experience-dependent plasticity on dendritic spines in the developing rat barrel cortex in vivo. Nature, vol. 404, No. 6780, 876-881.
Liu, D., Diorio, J., Tannenbaum, B., Caldji, C.,
Melchner, L., Pallas, S., Sur, M. (2000). Visual behaviour mediated by retinal projections directed to the auditory pathway. Nature, vol. 404, no. 6780 (20 de abril), p. 871-875.
Merzenich, M. (2000). Seeing in the sound zone. Nature, vol. 404, p. 820.
---, Neiderhiser, J., Hetherington, E. M. & Plomin, R. (2000). The Relationship Code. Cambridge, MA: Harvard University Press.
Neubauer, P. B. (1996). Nature's Thumbprint. 2nd ed. New York: Columbia University Press.
Plomin, R., DeFries, J. C., McLearn, G. E. & Rutter, R. (1997). Behavioral Genetics, 3rd ed. New York: Freeman.
Plotsky, P. M. & Meaney, M. J. (1993). Early, postnatal experience alters hypothalamic corticotropin-releasing factor (CRF) mRNA, median eminence CRF content and stress-induced release in adult rats. Brain Res.: Mol. Brain Res., 18:195-200.
Plotsky, P. M., Levine, L. J., Wright. E. L., Dennis, W. R. & Newcomb, R. L. (1997). Music training causes long-term enhancement of preschool children´s spatial-temporal reasoning. Neurol, Res., 19: 2-8.
Rauschecker, J.P. (1995). Compensatory plasticity and sensory substitution in cerebral cortex. Trends Neurosc., vol 18, p. 36-43
Reiss, D., Neiderhiser, J., Hetherington, EM., Plomin, R. (2000). The Relationship Code: Deciphering Genetic and Social Patterns in Adolescent Development. Cambridge, MA: Harvard University Press.
Reiss, D., Hetherington, E. M., Plomin, R., Howe, G. W., Simmens, S. J., Henderson, S. H., O'Connor, T. J., Bussell, D. A., Anderson, E. R. & Law, T. (1995). Genetic questions for environmental studies: Differential parenting and psychopathology in adolescence. Arch. Gen. Psychiat., 52:925-936.
Rowe, D. (1994). The Limits of Family Influence. Genes, Experience and Behavior. New York: Guilford Press.
Sadato, N., Pascual-Leone, A., Grafman, J. et. al. (1996). Activation of the primary visual cortex by reading in blind subjects. Nature, vol. 380: 526-528.
Sharma, J., Angelucci, A., Sur, M. (2000). Induction of visual orientation modules in auditory cortex. Nature, vol. 404, no. 6780 (20 de abril), p. 841-847.
Tienari, P., Wynne, L. C., Moring, J., Lahti, I. & Naarala, M. (1994). The Finnish adoptive family study of schizophrenia: Implications for family research. Brit. J. Psychiat., 23(Suppl. 164):20-26.
Tronick, E. (1989). Infant-mother face-to-face interaction: Age and gender differences in coordination and miscoordination. Child Development, 59, 85-92.
Zajonc, R. (1985). Emotion and facial efference: A theory reclaimed. Science, 228: 15-22.
Sponsored Links : Freshco Flyer, Giant Tiger Flyer, Loblaws Flyer, Kaufland Prospekt, Netto Marken-Discount Angebote